

En general, el color del plumaje de las aves viene determinado por la presencia de pigmentos como las melaninas y los carotenoides, por la concentración de dichos pigmentos en las plumas, por la naturaleza de la estructura queratinosa que forma las barbas y las bárbulas que conforman las plumas y por otros factores inherentes a la dotación genética de cada individuo, como son las hormonas o las enzimas (proteínas encargadas de aumentar la velocidad con la que se llevan a cabo infinidad de reacciones químicas en los organismos) implicadas en la síntesis y modificación de pigmentos. Así por ejemplo, un pájaro que se alimenta con semillas ricas en carotenoides y cuyas plumas son normales desde un punto de vista estructural, puede ser totalmente albino si, debido a mutaciones en su material genético, se suprimen todas las reacciones y mecanismos químicos implicados en la síntesis de melaninas y en la asimilación de carotenoides.
Al contrario de lo que ocurre con las melaninas, y salvo algunas excepciones, los animales no son capaces de sintetizar carotenoides sino que deben adquirirlos a través de la dieta. En muchos casos, los carotenoides asimilados se depositan tal cual en estructuras como las plumas o los caparazones de animales; no obstante, otros carotenoides pueden ser modificados por el animal mediante diversas reacciones químicas, que tienen lugar durante los procesos de digestión y fijación de los pigmentos, de manera que el pigmento que finalmente es fijado es un compuesto químico diferente. Este hecho, observado en infinidad de especies de aves, sugiere que, en gran medida, la coloración final de las plumas depende de la dotación genética de los individuos, la cual hace posible el desarrollo de estas reacciones químicas. Asimismo sugiere que la coloración final queda influida por la presencia de carotenoides en los alimentos de las aves.
El siguiente artículo supone una exposición y análisis de los factores que hacen posible la manifestación del color amarillo en las plumas de las aves de pequeño porte, haciendo especial hincapié en los canarios. Entre los factores determinantes de dicho color, se establecen, como principales, la dotación genética de los ejemplares y el sexo, así como la naturaleza y concentración de los pigmentos ingeridos por los canarios a través de la dieta. Entre dichos pigmentos cabe destacar aquellos conocidos como xantofilas, pigmentos de origen vegetal caracterizados por tener en su molécula átomos de carbono, hidrógeno y oxígeno.
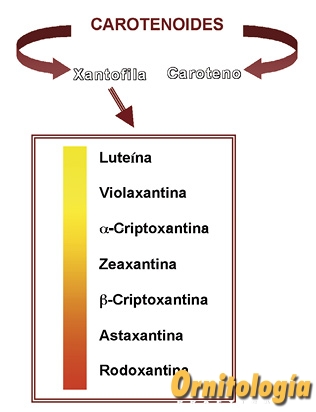
FIG.1 Esquema resumen de la clasificación de los carotenoides según su composición química, destacando las xantofilas más abundantes en la naturaleza.
Mientras que algunas xantofilas son ingeridas, asimiladas y fijadas en las plumas de las aves sin sufrir ningún cambio químico, otras experimentan modificaciones en su composición química, tal y como se citó anteriormente. Los productos finales así obtenidos tienen diferentes propiedades de absorción de luz, de ahí que se manifiesten colores con distinta propiedad. Así, es posible observar una amplia variedad de colores en las plumas e incluso diferentes tonalidades para un mismo color.
Índice del artículo
Carotenoides en la naturaleza
Los carotenoides son pigmentos con tonalidades que van desde el amarillo al púrpura, pasando por toda una gama de rojos y anaranjados. Estos pigmentos son sintetizados principalmente por vegetales y algunas algas; aunque ciertos hongos, levaduras y bacterias también son capaces de producirlos. Los animales adquieren los carotenoides a través de la alimentación, puesto que no los pueden sintetizar.

Escribano de Brandt
De forma general, los carotenoides se clasifican en dos grandes grupos: los carotenoides oxigenados (tienen oxígeno en su estructura), denominados xantofilas, y los no oxigenados (ausencia de oxígeno en la estructura), que reciben el nombre genérico de carotenos: el beta-caroteno (b-caroteno) y sus derivados (Fig. 1).
Las xantofilas aparecen en la naturaleza como pigmentos amarillos, pardos o rojizos. La intensidad de color de las mismas varía dependiendo del número de átomos de oxígeno presentes en la molécula. Las xantofilas que se han caracterizado hasta la fecha, se han obtenido en la mayor parte de los casos de vegetales. No obstante, a medida que se estudia con detalle la coloración de los animales, en especial las aves, se han detectado nuevas variedades de xantofilas, que son en realidad xantofilas ingeridas por los animales y posteriormente modificadas en el organismo antes de que se depositen en estructuras como las plumas.
Color amarillo en aves
Tal y como se ha mencionado en el apartado anterior, los carotenoides ingeridos pueden ser incorporados en el cuerpo del animal tal cual o pueden ser modificados, gracias a diversas reacciones químicas, antes de que se depositen en determinadas estructuras o tejidos del cuerpo.

Canario Satiné amarillo intenso.
Los mecanismos genéticos y enzimáticos que controlan el metabolismo de los carotenoides y su fijación en las plumas en aves no están, por el momento, bien establecidos. En los últimos 10 años aproximadamente, científicos que trabajan en este campo se han formulado una serie de preguntas que podrían ser resumidas en las siguientes:
1. ¿Todas las aves asimilan por igual todos los carotenoides?
2. ¿Se acumulan por igual en todos los tejidos de los animales los carotenoides ingeridos?
3. El color amarillo que se observa, ¿es sólo debido a los carotenoides?
4. ¿Todos lo carotenoides contribuyen a la generación de tonalidades intensas en las aves?
5. Independientemente del efecto que tiene el color estructural ¿la cantidad y variedad de carotenoides ingeridos determina el color final del animal?
La mayor parte de estas incógnitas siguen sin estar resueltas de forma precisa, aunque poco a poco se va teniendo más detalle gracias a estudios realizados con jilgueros, cardenalitos y pollos, básicamente. Como resultado de estos estudios, y en respuesta a las preguntas anteriores, por el momento sabemos que:
1. Todas las aves no asimilan y fijan todos los carotenoides por igual. Parece que de todos los carotenoides presentes en las semillas que las aves ingieren, la luteína y la zeaxantina son los que de forma mayoritaria asimilan la mayor parte de las aves estudiadas. No obstante, dependiendo de las especies, incluso la zeaxantina es preferentemente asimilada y fijada.
2. Los pigmentos no se acumulan por igual en todos los tejidos y estructuras de los animales y cada pigmento proporciona un color particular a dichas estructuras, teniendo en cuenta que en muchos casos el color final que observamos se debe a una mezcla de pigmentos en el mismo tejido o estructura. Ejemplos de esta distribución variada de pigmentos según zonas la constituyen los jilgueros y los pinzones, entre otros. En el caso de los pinzones, por ejemplo, el color amarillo-verde de la mayor parte del cuerpo es debido a la fijación de la luteína tal cual se obtiene de los alimentos, quedando finalmente mezclada con las melaninas.
3. El color amarillo final de los animales, y en particular de las plumas de las aves, no sólo depende de la cantidad y variedad de pigmentos presentes (aunque este factor es muy importante) sino que también está determinado por la naturaleza de la estructura queratinosa de las plumas.
4. No todos los carotenoides contribuyen a la generación de tonalidades intensas en las plumas. Experimentos realizados con machos de jilgueros americanos (Carduelis tristis) con una dieta suplementada sólo con zeaxantina o sólo con luteína pusieron de manifiesto que los machos tenían tonalidades más intensas cuando se alimentaban con zeaxantina. No obstante, cabe resaltar que en estas aves la zeaxantina no se fija como tal, sino que es modificada antes de ser depositada en las plumas.
5. En general, se puede decir que sí se cumple en la mayoría de las aves que a mayor cantidad de carotenoides ingeridos, más intensa es la coloración del plumaje. Sin embargo, en estudios realizados con jilgueros americanos machos y hembras, se ha observado que cuando todos los ejemplares (tanto machos como hembras) eran sometidos a la misma dieta en ausencia o presencia de pigmentos, desarrollaban distinto color, siendo siempre los machos los que mayor intensidad de color tienen. Esto pone de manifiesto que la dotación genética de los individuos, sistema hormonal, etc., determina en gran medida el color final. Este no es un ejemplo aislado sino que existen muchas especies en las que se cumple este patrón.

Canario Opal amarillo nevado.
Hasta hace relativamente poco, expertos y aficionados consideraban que los colores rojo-amarillo finales de las aves se debían a los carotenoides presentes en las semillas, los cuales “son fijados tal cual” en las plumas, una vez ingeridos. Partiendo de esta idea se ha extendido la práctica de aumentar en la dieta de las aves criadas en cautividad la dosis de aquellas semillas con mayor contenido en carotenoides. Otra alternativa, muy usada en sectores como la canaricultura, es suministrar en la pasta de cría compuestos químicos a base de carotenoides (y concretamente xantofilas, casi siempre) de gran pureza. No obstante, se sabe que en muchas especies, los carotenoides que están presentes en las plumas, no son los presentes en las semillas ingeridas, sino que son compuestos químicos derivados de los mismos. Por el momento, se han caracterizado algunos carotenoides modificados aislados directamente de las plumas y entre ellos, por su abundancia, se pueden destacar un conjunto de xantofilas denominadas -caroten-3-onas.
Xantofilas del Canario
Como se ha indicado anteriormente, todos los estudios apuntan a que la luteína y la zeaxantina son los pigmentos más abundantes en las plumas y los que actúan como precursores de otras variantes que se localizan también en las plumas. En este sentido, el canario no es una excepción.
Dentro el conjunto de pigmentos que aparecen en las aves por modificación de los presentes en la dieta, se encuentra un grupo de compuestos denominados -caroten-3-ones constituido por xantofilas como la 3-dehidroluteina, la xantofila A del canario y la xantofila B del canario, pigmentos amarillos encargados de proporcionar color amarillo brillante al plumaje. Algunos artículos de divulgación se han hecho eco de la existencia de una tercera y cuarta xantofila del canario, conocidas como xantofila del canario C y xantofila del canario D. Lo cierto es que no hay muchas publicaciones ni estudios rigurosos sobre la existencia o la naturaleza de estas dos últimas xantofilas del canario. El hecho de que las xantofilas de canario A y B reciban ese nombre no se debe a que sean exclusivas de los canarios, de hecho estos pigmentos también se han aislado de plumas de cardenalitos con coloración amarilla aberrante.
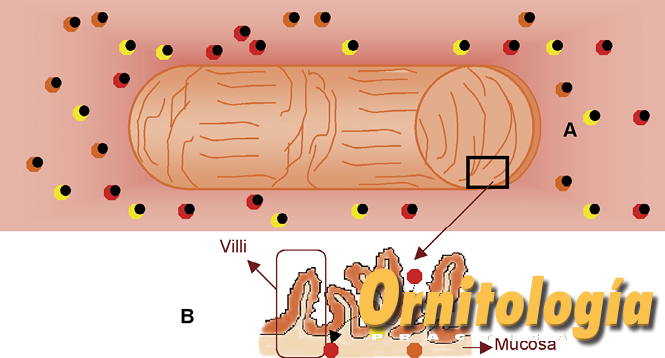
Figura 3
Según las observaciones realizadas con ejemplares de saltarín militar (Ilicura militaris), cardenal rojo (Cardinalis cardinalis) o canario, se ha propuesto que la xantofila del canario tipo A se sintetiza en los pájaros a partir de la luteína de la dieta. Gracias a determinadas reacciones químicas, la xantofila del canario A puede seguir siendo modificada para dar lugar a la xantofila B, aunque esta última se puede obtener directamente por modificación de la zeaxantina. (Fig.2).
Fijación de las xantofilas en las plumas
Cuando las aves ingieren las semillas, se desencadena toda una serie de reacciones químicas propias del proceso de digestión, lo que permite que finalmente los animales obtengan nutrientes de los alimentos tales como las vitaminas, sales minerales, proteínas, etc. Gracias a este proceso, las aves obtienen también los carotenoides presentes en la dieta. Dichos carotenoides pueden ser modificados o no por el animal, tal como se ha descrito. En cualquiera de los casos, los carotenoides deben ser absorbidos, en el intestino delgado, principalmente. Con objeto de facilitar dicha absorción, la mucosa del intestino posee una serie de proyecciones, como dedos de guante, llamadas villi o vellosidades. En el interior de cada villi existe una vénula, una arteriola y un vaso linfático, que recogerán los carotenoides absorbidos y los distribuirán a lo largo de todo el organismo, gracias al torrente sanguíneo (Fig. 3).
El transporte de las xantofilas a través de la sangre, a priori es un problema, puesto que las xantofilas son compuestos liposolubles, es decir son solubles en medios grasos pero no en agua. Teniendo en cuenta que la sangre, en su mayor parte, está constituida por agua, según lo comentado, sería imposible que las xantofilas se mantuvieran solubles en la sangre y por tanto su transporte a las distintas partes del cuerpo del animal sería casi imposible. Este problema se solventa en los animales gracias a la unión de las xantofilas a unas proteínas denominadas lipoproteínas, que actúan como “facilitadores del transporte” de las xantofilas a través de la sangre. Es por esto que muchos criadores de canarios suministran carotenoides en la dieta junto con pequeñas dosis de aceite, para favorecer la asimilación de los pigmentos. Aunque esto es una práctica habitual en los criaderos, no se ha cuantificado ni analizado con exactitud si la adición de aceite mejora o no la asimilación de los pigmentos con los que se suplementa la dieta de los canarios.

Figura 4
La naturaleza liposoluble de los carotenoides hace posible que mayoritariamente estos pigmentos se acumulen en regiones grasas de los pájaros tales como el tejido adiposo (tejido constituido por unas células denominadas adipocitos, que son la reserva de grasas en animales) o en el hígado.
Una vez que las xantofilas se encuentran asociadas a lipoproteínas en sangre, éstas son transportadas por todo el cuerpo, para llegar a las regiones en las que han de fijarse, incluyendo las plumas. Las plumas crecen en papilas o folículos especializados de la piel. Estos folículos son similares a aquellos en los que crece el pelo en los mamíferos y tienen en su base músculos y terminaciones nerviosas. A diferencia de lo que podríamos pensar, las plumas no crecen en todo el cuerpo, sino que lo hacen en zonas particulares llamadas pterilos (áreas donde se encuentran las papilas que dan origen a las plumas nuevas). Estos pterilos tienen una forma y disposición particular para cada especie, y están rodeados por zonas desnudas llamadas apterios, en las que no crecen plumas.
La estructura de una pluma es muy compleja; se trata de una lámina compuesta por gran cantidad de elementos (Fig. 4)
En una pluma típica, la parte central (raquis) sirve de eje y tiene el aspecto de una caña hueca; a pesar de ser una estructura muy ligera, confiere a la pluma la rigidez necesaria para mantenerla firme. La parte inferior del raquis, más ancha, hueca y generalmente desnuda, se denomina cálamo o cañón. Mediante el cálamo, la pluma se inserta en la piel. En la parte inferior del cálamo hay un orificio, denominado ombligo inferior, que es por donde la pluma se alimenta durante su crecimiento. El cañón es hueco, puesto que la pluma es una estructura muerta, como las uñas, y una vez que ha terminado de crecer, los vasos sanguíneos que la alimentaban se retiran y queda el haz vacío.
Si se analiza la estructura de la pluma aquí expuesta, resulta fácil comprender el proceso de pigmentación de la misma mediante carotenoides. Cuando los carotenoides son absorbidos por el canario, el torrente sanguíneo los conduce hasta la epidermis. En aquellas zonas de la epidermis donde haya folículos, emerge el ombligo inferior y el cálamo, por cuyo interior circulan pequeños vasos sanguíneos mientras la pluma crece. Así, los carotenoides que llegan hasta allí se fijan en la pluma en formación (entre las láminas de queratina que conforman las plumas y en el raquis). Este proceso sólo ocurre mientras que la pluma está en formación (aproximadamente alrededor de los primeros 30-40 días de vida de los polluelos o durante la muda), puesto que sólo en este momento existen vasos sanguíneos en su estructura gracias a los cuales llegan los pigmentos.
Fijación de las xantofilas en las yemas de los huevos
Durante la formación de los huevos, y en particular de la yema, el organismo emplea las reservas de carotenoides que se encuentran acumuladas en el tejido adiposo y en hígado. De nuevo gracias a la conducción de los carotenoides a través del torrente sanguíneo, éstos se movilizan para llegar hasta la yema en formación. Si las reservas de carotenoides son escasas, el color final de la yema será pálido.
En la cría de aves en cautividad, este problema se ha resuelto mediante el suministro de pastas de alto contenido en carotenoides. Ahora bien, para obtener buenos resultados es imprescindible llevar a cabo una buena aplicación de las mismas. Si dichas pastas se proporcionan a las aves cuando ya ha comenzado la puesta, las primeras puestas darán lugar a huevos de yemas pálidas. Este problema degenera en otro mayor si cabe: los nidáceas asimilan los carotenoides de la yema durante un mes aproximadamente, por lo que si la yema era deficitaria en carotenoides, las nidáceas no asimilarán de forma óptima la mayor cantidad de carotenoides posible, lo que dará lugar a ejemplares de color pálido, resultado nefasto en el caso de que se trate de crías de canarios de factor rojo, por ejemplo.
Nuevas tendencias en el estudio de las xantofilas en canarios
Durante siglos, buena parte del trabajo y de las mejoras que se han realizado en el control de la pigmentación de aves de pequeño porte han tenido lugar gracias a la pasión y entusiasmo mostrado por infinidad de criadores. Mediante la observación y el establecimiento de cruces, se ha desarrollado toda la disciplina de genética aplicada en muchos casos, aún en perfecto desconocimiento de las bases teóricas de estos procesos, lo cual es merecedor del máximo reconocimiento.
-


- Las barbas y las bárbulas de estas plumas de la región cervical presentan una distribución del lipocromo que se extiende en toda su longitud.
-


- A la derecha, la pluma de ejemplar nevado, en el que, en la región del cuello, la nevadura apenas se insinua.
Actualmente, numerosos grupos de investigación están intentando demostrar y explicar de forma más objetiva y cuantificable, cómo se asimilan los carotenoides, cómo se fijan en los tejidos, qué reacciones hacen posible la obtención en el animal de otros carotenoides, qué diferencias hay entre los carotenoides transportados en sangre y los que hay en las plumas, etc. En relación con este punto, lo que se está haciendo actualmente es trabajar con muestras de sangre y plumas recogidas de diversos ejemplares de distintas especies (jilgueros principalmente) para llevar a cabo extracciones directas de pigmentos. La muestra que resulta de dichas extracciones es analizada con una técnica propia de la Bioquímica denominada Cromatografía líquida de alta presión (HPLC), la cual permite separar distintos compuestos de una mezcla. Por tanto, teniendo varios carotenoides en una mezcla, estos se pueden separar por HPLC, para luego ser analizados químicamente por separado, haciendo uso de otras técnicas como la espectrofotometría.
Para la mayor parte de los criadores, esta nueva forma de análisis queda fuera de su alcance en muchos sentidos, mientras que para la mayor parte de científicos que analizan estos procesos quedan fuera de su conocimiento los aspectos más básicos de cría y cruce de las aves según la genética más tradicional (aspecto mejor controlado por los criadores). Sería interesante fomentar foros de encuentro y discusión de ambos sectores para contribuir al conocimiento de una temática tan apasionante como es el color de las aves y su papel en la naturaleza.
Conclusiones
• Los carotenoides luteína (en mayor medida) y zeaxantina son los precursores de las xantofilas conocidas como Xantofilas del canario.
• Las Xantofilas del canario, presentes también en otras aves, proporcionan colores amarillos de considerable brillo.
• En general, luteína y zeaxantina dan lugar a la síntesis de prácticamente la totalidad de pigmentos amarillos existentes en aves.
• Luteína y zeaxantina, en especial el primero, son abundantes en semillas y vegetales. No obstante, también es posible encontrarlos en grandes concentraciones en las larvas de algunos insectos, los cuales forman parte de la dieta de muchas aves en libertad.
• De lo anterior se deduce que aquellos alimentos ricos en luteína favorecerán una expresión máxima del color amarillo en el plumaje, especialmente en los ejemplares genéticamente bien dotados. Es importante realizar una buena aplicación de las pastas de cría ricas en carotenoides para obtener los mejores resultados posibles.
Bibliografía
McGraw KJ, Hill GE, Navara KJ, Parker RS (2004) Differential accumulation and pigmenting ability of dietary carotenoids in colourful finches. Physiol. Biochem. Zool. 77: 484-491.
Hudon J, Anciães M, Bertacche V, Stradi R (2007) Plumage carotenoids of the Pin-tailed Manakin (Ilicura militaris): Evidence for the endogenous production of rhodoxanthin from a colour variant. Comparative Biochemistry and Physiology, Part B 147:402–411.
Wolf P, Bartels T, Sallmann HP, Heisler K, Kamphues J (2000) Vitamin A metabolism in recessive white canaries. Animal-Welfare 9:153-165.


